Isolamento das ilhas florestais é o principal fator que influencia a mudança temporal da composição de insetos nos capões de mata da Serra do Cipó
Ilhas já foram alvo de interesse de vários cientistas no passado e continuam atraindo a atenção até os tempos atuais. Pesquisadores interessados em História Natural, Genética, Zoologia, Botânica e Ecologia se interessam por ilhas desde o século XVIII. Essas ideias foram melhoradas e culminaram no livro Biogeografia de Ilhas (1967), de autoria de um matemático Robert MacArthur e um biólogo e evolucionista Edward Wilson, que causou uma revolução nos pensamentos da época e se tornou um clássico. A Teoria de Biogeografia de Ilhas (TBGI) examina padrões de biodiversidade em ilhas e fornece as principais ideias e hipóteses sobre como o isolamento e o tamanho da ilha afetam a dinâmica das populações e comunidades. Por exemplo, o equilíbrio entre a taxa de colonização (ou chegada) de novas espécies e a taxa de extinção (ou desaparecimento) de espécies existentes é um aspecto fundamental da dinâmica de populações e comunidades em ilhas.

Na ecologia moderna, o termo “ilhas” não está restrito somente às ilhas oceânicas, como proposto inicialmente por MacArthur e Wilson. Na ecologia o termo “ilhas” é mais amplo e geralmente se refere a áreas de habitat isoladas e distintas, cercadas por um ambiente diferente que impede ou dificulta a movimentação de organismos de uma ilha para outra. Dentro dessa ideias, “ilhas” podem incluir, por exemplo, fragmentos de florestas isolados em uma paisagem agrícola ou até mesmo pequenos corpos d’água que são isolados por características geográficas ou climáticas. A ecologia de ilhas é uma área de estudo que examina os padrões e processos que ocorrem em ambientes fragmentados naturalmente – caso dos capões de mata (Figura 1 e 2) – ou de forma antropogênica – quando uma floresta é dividida em fragmentos menores, causando a fragmentação e perda de habitat. Alguns dos conceitos-chave da TBGI de MacArthur e Wilson incluem o isolamento da ilha, o tamanho das ilhas, a taxa de colonização e de extinção de espécies nessas ilhas. O isolamento se refere à distância física entre as ilhas e o continente, ou manchas de habitat mais extensas, conhecidas como áreas doadoras de espécies. Ilhas mais isoladas tendem a ter menos intercâmbio de espécies com outras ilhas ao longo do tempo devido à limitação de dispersão presente em várias espécies. Logo, ilhas florestais mais distantes são esperadas terem uma composição mais diferente do continente (ou da área doadora) se comparadas às ilhas mais próximas, umas vez que ilhas mais distantes serão difíceis de serem alcançadas, o que influencia diretamente na taxa de colonização de espécies nessas ilhas. O tamanho da ilha é outro componente chave na TBGI. Em geral, ilhas maiores têm maior capacidade de suportar uma variedade (heterogeneidade) de habitats e também de populações maiores, que determinam menores perdas ou taxas de extinção das espécies. Isso pode se refletir em uma maior riqueza de espécies presente em ilhas florestais maiores do que em ilhas menores. Pensando em ilhas florestais, hipóteses mais recentes apontam que a quantidade (ou área) de floresta em uma paisagem fragmentada pode ter um efeito ainda maior do que o isolamento e o tamanho das ilhas juntos. Em outras palavras, uma maior quantidade de manchas de florestas (ou ilhas) juntas pode ter efeito positivo na riqueza de espécies, independentemente do isolamento ou do tamanho de cada ilha. Isso quer dizer que as características individuais de cada ilha podem ser menos importantes e que uma abordagem em nível de paisagem pode ser mais adequada para se entender melhor os padrões encontrados em cada ilha florestal. Portanto, a diversidade de espécies em uma ilha florestal pode ser influenciada pelo isolamento, tamanho da ilha, quantidade de habitat na paisagem e outros fatores ambientais, como a disponibilidade de alimento e de condições diferentes entre as ilhas (p.ex. dossel mais ou menos aberto em diferentes capões de mata).

Atualmente, a TBGI faz parte do arcabouço teórico que lança mão de ideias de metacomunidades, que são comunidades conectadas pela dispersão de espécies ou indivíduos entre ilhas (ou manchas de habitat). Em metacomunidades, há teorias que levam em consideração o papel da dispersão das espécies, características do habitat e interações, que são fundamentais para a manutenção de espécies nas manchas (ou ilhas) e, assim, da diversidade local e regional das comunidades biológicas. Porém, ainda não se sabe muito como grupos com diferentes capacidades de dispersão – como é o caso dos insetos – respondem às características das ilhas florestais na paisagem (isolamento e tamanho) em termos de mudanças na composição ao longo do tempo (diversidade beta temporal). A mudança na composição ao longo do tempo se dá basicamente por dois processos: colonização (chegada de espécies diferentes das já presentes) e extinção (desaparecimento local das espécies).
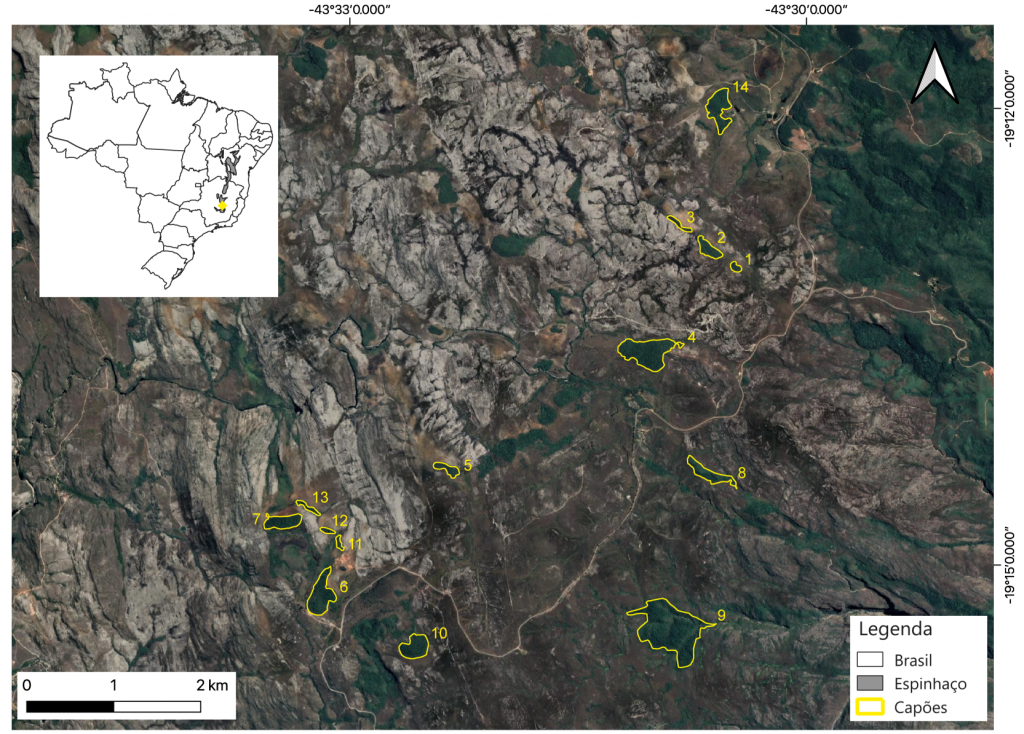
Nas regiões mais altas da Cadeia do Espinhaço (acima de mil metros de elevação), montanhas que nascem no meio do estado do Minas Gerais e se expendem até o meio da Bahia, são comum ilhas florestais conhecidas como os “capões de mata” (Figura 1 e 2), que formam verdadeiros arquipélagos florestais imersos em uma matriz de campos rupestres, os quais podem funcionar como barreiras para espécies que são fortemente associadas às condições ou aos recursos fornecidos exclusivamente pelas ilhas de floresta. Com o intuito de entender melhor os processos que geram os padrões atuais de diversidade ao longo do tempo de cinco grupos de insetos, nós avaliamos os efeitos do tamanho da ilha florestal, do isolamento e da área de floresta na paisagem que impulsionam mudanças temporais, incluindo ganhos e perdas de biodiversidade de insetos em um arquipélago florestal no topo da Serra do Cipó. Nós esperávamos que em ilhas florestais menores e mais isoladas, as mudanças na composição dos insetos fossem mais proeminentes, impulsionadas principalmente por ganhos de espécies ao longo do tempo. Além disso, a maior quantidade de floresta na paisagem levaria a maiores ganhos de espécies boas dispersoras ao longo do tempo, independentemente do tamanho da ilha florestal e do isolamento. Por fim, nós esperávamos que grupos de insetos com dispersão mais limitada sofressem maior diferenciação ao longo do tempo, enquanto em grupos com elevada capacidade de dispersão teríamos uma maior homogeneização de espécies entre as ilhas, ou seja, as mesmas espécies de elevada capacidade de dispersão seriam encontradas em diferentes ilhas florestais. Para testar essas hipóteses, nós amostramos diferentes grupos de insetos para representar um “gradiente de dispersão”. Utilizamos os seguintes grupos: formigas, rola-bostas, abelhas, vespas e borboletas frugívoras, sendo as formigas o grupo com menor capacidade de dispersão, pois a biologia da maioria das espécies envolve períodos de indivíduos com asas em um curto período de tempo (período de acasalamento e formação de uma nova colônia). No outro extremo, as borboletas são tidas como tendo alta capacidade de dispersão, onde algumas espécies podem migrar por centenas de quilômetros. Os demais grupos – rola-bostas, abelhas e vespas – são tidos como intermediários em capacidade de dispersão, quando comparados com formigas e borboletas. Esses grupos foram amostrados na estação chuvosa com métodos padronizados de coleta para cada grupo em 14 capões de mata de diferentes dimensões em 2014, 2015, 2018 e 2020 (Figura 3).
Como principais resultados, nós encontramos que processos distintos de homogeneização e heterogeneização (ou diferenciação) resultantes de colonização (chegada) e extinção (desaparecimento) estão agindo de acordo com cada grupo de insetos, provavelmente devido a diferentes capacidades de dispersão. Por exemplo, as perdas de espécies predominaram nas formigas, com espécies raras e espécies mais comuns sendo perdidas ao longo dos anos de amostragem. As borboletas ganharam espécies ao longo do período de amostragem, representadas principalmente por espécies mais comuns, levando a um aumento da homogeneização resultante da colonização de espécies que são comumente encontradas. A distância entre ilhas florestais vizinhas (isolamento) foi o principal fator que afetou a mudança de composição temporal dos grupos de insetos, determinando também os ganhos e perdas de espécies (Figura 4).
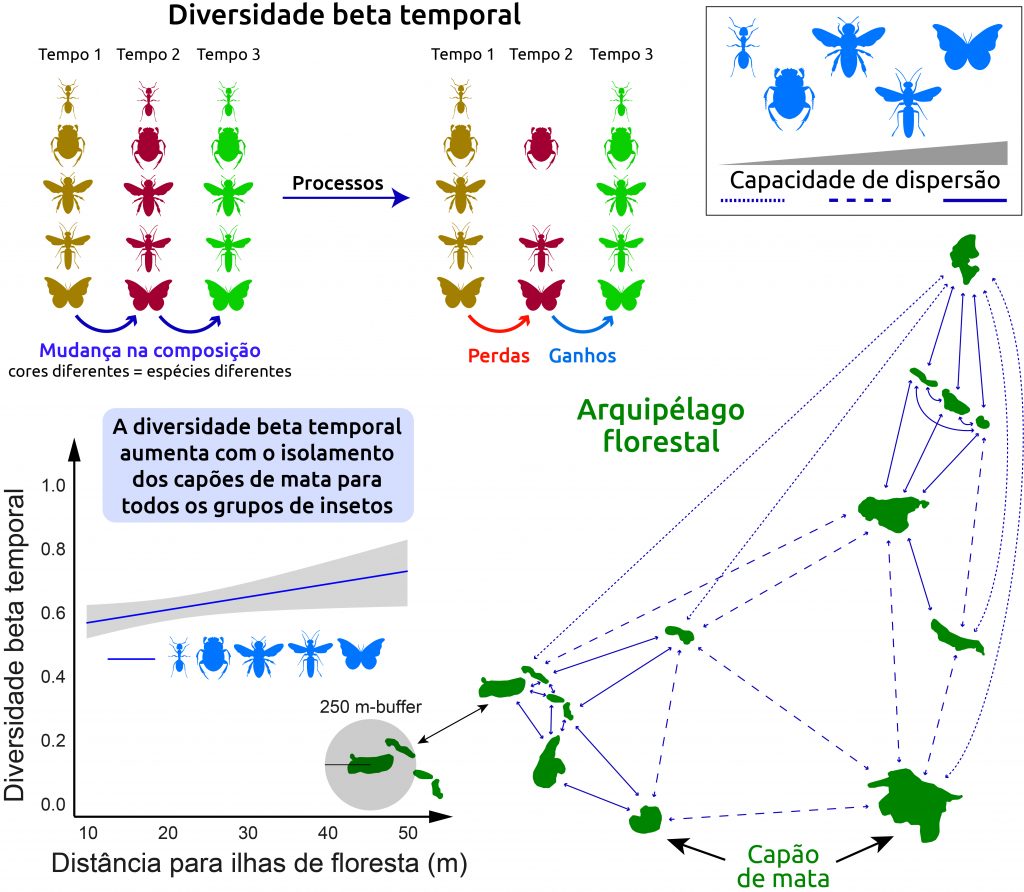
Essas descobertas fornecem informações biogeográficas valiosas sobre as interações complexas entre as características das ilhas florestais, os atributos da paisagem e as capacidades de dispersão que moldam a dinâmica temporal da biodiversidade de insetos nos topos das montanhas. Assim, conservar a quantidade de floresta na paisagem e manter a conectividade florestal entre os capões de mata são necessários porque a dinâmica temporal de colonização e desaparecimento local das espécies de insetos pode depender da capacidade de dispersão desses organismos.
Autores
Pedro Giovâni da Silva
Universidade de Brasília, Instituto de Ciências Biológicas, Departamento de Ecologia, 70910-900, Brasília, DF. pedrogiovanidasilva@yahoo.com.br
Frederico de Siqueira Neves
Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas, Departamento de Genética, Ecologia e Evolução, 31270-901, Belo Horizonte, MG. fred.neves@gmail.com
Fonte
Da Silva, P. G.; Beirão, M. V.; Castro, F. S.; Perillo, L. N.; Camarota, F.; Solar, R. R. C.; Fernandes, G. W.; Neves, F. S. 2023. Isolation drives species gains and losses of insect metacommunities over time in a mountaintop forest archipelago. Journal of Biogeography 50(12): 2069-2083. https://doi.org/10.1111/jbi.14714